疏水性氨基酸和亲水性氨基酸的0
生物学家劳伦斯·赫斯特和斯蒂芬·弗里兰在20世纪90年代末把天然基因密码和计算机随机产生的几百万组密码拿去比对,结果轰动一时。他们想知道,如果发生点突变这种把一个字母换掉的变异,哪一套密码系统能保留最多正确的氨基酸,或将它代换成另一个性质相似的氨基酸。
结果他们发现,天然的基因密码最经得起突变的考验。点突变常常不会影响氨基酸序列,而如果突变真的改变了氨基酸,也会由另一个物理特性相似的氨基酸来取代。据此,赫斯特与弗里兰宣称,天然的遗传密码比成千上万套随机产生的密码要优良得多。它不但不是大自然密码学家愚蠢而盲目的作品,而是万里挑一的密码系统。
天然的三联基因密码的第一个字母都有特定的对应方式。举例来说,所有以丙酮酸为前体合成的氨基酸,它们密码的第一个字母都是T。所有由α-酮戊二酸所合成的氨基酸,其三联密码第一个字母都是C;所有由草酰乙酸合成的氨基酸,第一个字母都是A;最后,几种简单前体通过单一步骤所合成的氨基酸,第一个字母都是G。
三联密码的第二个字母和氨基酸是否容易溶于水有关,或者说和氨基酸的疏水性有关。亲水性氨基酸会溶于水,疏水性氨基酸不会溶于水,但会溶在脂肪或油里,比如溶在含有脂质的细胞膜里。所有的氨基酸,可以从“非常疏水”到“非常亲水”排列成一张图谱,而正是这张图谱决定了氨基酸与第二个密码字母之间的关系。疏水性最强的六个氨基酸里有五个,第二个字母都是T,所有亲水性最强的氨基酸第二个字母都是A。介于中间的有些是G有些是C。
三联密码的第三个字母不含任何信息,不管接上哪一个字母都没关系,这组密码子都会翻译出一样的氨基酸。以甘氨酸为例,它的密码子是GGG,但是最后一个G可以代换成T、A或C。
第三个字母的随机性暗示了一些有趣的事情。二联密码可以编码16种氨基酸。如果我们从20个氨基酸里拿掉5个结构最复杂的(剩下15个氨基酸,再加上一个终止密码子)这样前两个字母与这15个氨基酸特性之间的关联就更明显了。因此,最原始的密码可能只是二联密码,后来才靠“密码子捕捉”的方式成为三联密码,也就是各氨基酸彼此竞争第三个字母。
第一个字母和氨基酸前体之间的关系直截了当,第二个字母和氨基酸的疏水性相关,第三个字母可以随机选择。这套密码系统除了可以忍受突变,还可以降低灾难发生时造成的损失,同时可以加快进化的脚步。因为如果突变不是灾难性的,那应该会带来更多的好处。
秸秆制塑料

植物纤维高分子复合仿木产品
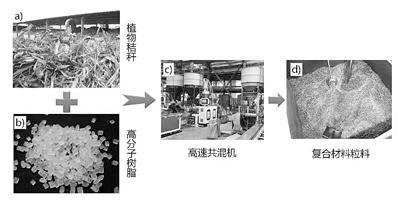
植物纤维高分子复合材料制备过程
2017年,中国人造板材产量超过3亿立方米,其中60%的原材料依赖进口。与此同时,各类秸秆可收集量超过8亿吨,其中只有极少部分得到有效再利用。研发植物纤维高分子复合材料,就是为了从根本上解决木材产需之间的矛盾。
“像木材一样可钉可铆,点燃10秒后离火自灭,不会翘曲也不发生霉变,而且价格比普通木材还要低……”近期,中科院宁波材料所研究员朱锦将草本植物纤维与塑料树脂结合,研制出了一种新型的仿木材料——植物纤维高分子复合材料,并且其对水火还具有“免疫”属性。
“以往的原木或仿木材料,在性能优势上往往顾此失彼,且离不开对木材的消耗。”朱锦告诉科技日报记者,这一材料的研制及相关技术的突破,将减少人类对木材的依赖,促进木材的可持续发展。目前,研究团队通过与一家山东科企合作,已能制备出植物纤维填充量高达80%的植物纤维高分子复合材料。
微观设计再现“古式纤维复合”
纤维沿着生长方向排列,形成了天然木材的各向异性——木材受压时,每个细胞都相当于一根管柱,在横纹、顺纹两种情况下会表现出不同的抗压能力,导致木材便于加工的同时也容易翘曲。“但我们所研制的复合材料因为拥有高含量且不规则的植物纤维,可保证材料在横纹、顺纹方向下都具有均匀的抗压性能,这使得复合材料不会翘曲,并且植物纤维高分子复合材料密度低,可锯、可铆、可钉、可刨,加工十分灵活。”朱锦说。
“纤维复合材料的历史可追溯至古代。古人将稻草或麦秸混入粘土增加粘土强度用以建造房屋,就是一种纤维复合材料的利用。”朱锦表示,由于秸秆难于收集处理,且易腐、易燃、易吸水,在现代复合材料中很难见到它们的身影。
但朱锦等人研制的植物纤维高分子复合材料,却用到了古人所用的秸秆。可制作这种材料仍需要解决几个问题。
首先,让高含量的植物纤维与塑料树脂进行充分共混,需要具有强力共混搅拌扭矩的硬件设备。“以往的木塑复合材料,以木粉作填充,填充量低于60%。而我们与山东科企合作研制的高速共混机,可通过其极强的共混搅拌扭矩,让高含量的植物纤维与塑料树脂进行充分共混,让微植物纤维填充量达80%。”朱锦解释道,这令它在复合材料中起到类似钢筋混凝土中“钢筋”的作用,让成品的力学性能远高于市场上的木塑复合材料。
其次,由于植物纤维具有较强的亲水性,其与疏水的高分子树脂之间相容性很差,会导制复合材料表现出很大的脆性。对此,朱锦表示,团队优选了合适的相容助剂,改善亲水植物纤维与疏水性高分子树脂之间的相容性,提高两者在微观层面的结合力,让植物纤维高分子复合材料具有了高强度、高模量。
最后还有个难点。植物纤维含量一旦提高,植物纤维高分子复合材料的熔体粘度(在一定温度下熔体流动性能的好坏,粘度越大,熔体流动性越差)也会相应增加,会使得材料在注塑过程中流动性变差,甚至无法充满模腔,最终获得的制件表面容易形成各式缺陷。为此,朱锦团队采用“内润滑”加“外润滑”的复配润滑剂体系,降低了熔体的内部粘度,减小了熔体与设备之间的摩擦。
独家配方增强仿木“体质”
众所周知,植物纤维与常见的塑料都是易燃品。植物纤维复合材料的阻燃问题依然是业界难题,很多研究者通过有机溶剂或者碱液处理纤维,破坏纤维表面的结构,然后加入阻燃剂进行阻燃。
“这种做法会产生大量的废弃溶剂和碱液,在环保处理方面增加了额外成本。”朱锦说,于是团队采用植物纤维与树脂基体分步阻燃化的方案,先用水对植物纤维进行简单的喷淋,再用常规的加工方法与环保型阻燃剂,使树脂获得足够的阻燃能力,植物纤维复合材料点燃10秒后,离火便自动熄灭。
防得了火,能否防潮?
实验团队将封闭样条浸泡在20℃的恒温水中,100天后,样条的吸水率仍然低于2%。为让样品多吸水,实验人员将挤出块材表面的塑料层切除,又进行了60天的吸水性测试,样品的吸水率均仍未超过20%,远低于30%的腐烂临界值。
“这是因为在高速共混机中,高分子基体与纤维达到了非常均匀的共混,完成了对植物纤维的有效包裹。”朱锦表示,团队做了大量精密计算,才得出配方设计,让高达80%植物纤维含量的复合材料依然能够受到高分子基体的保护,大幅度降低吸水性能。同时保证了植物纤维复合材料在高温湿热的情况下,依然具有良好的稳定性能。
“目前团队已经贯通了植物纤维高分子复合材料的整体工艺路线,具备了一定的生产与制备能力,并使产品在成本与使用性上取得了良好的平衡。”朱锦表示,随着研发和市场推广的深入,这款仿木产品会很快进入市场,从而促成人类将树木还给森林。(本文图片由受访单位提供)
跟着塑料“混” 秸秆变身“木材”水火不侵,秸秆制塑料
《原清华大学生物学教授颜宁在科学技术实验上的探索与创新》
1996年-2000年清华大学生物科学与技术系学士;
2000年-2004年美国普林斯顿大学分子生物学系,博士,导师为结构生物学家、清华大学教授、中国科学院院士、欧洲分子生物学学会外籍会士、美国国家科学院外籍院士、美国人文与科学院外籍院士施一公;
2005年-2007年 美国普林斯顿大学分子生物学系从事博士后研究;
2007年-至今清华大学教授、博士生导师;
2017年5月7日从清华大学证实,颜宁已接受美国普林斯顿大学邀请,受聘该校分子生物学系雪莉·蒂尔曼终身讲席教授的职位。
研究方向
人类基因组中编码蛋白的所有基因约有30%编码膜蛋白。
膜蛋白在一切生命过程中起着关键作用,具有重要的生理功能。FDA批准上市的药物中,约50%的作用靶点为膜蛋白。
因此,对膜蛋白结构与功能的研究具有极高的生物学意义及医药应用前景。
转运蛋白(transport proteins)是膜蛋白的一大类,介导生物膜内外的化学物质以及信号交换。脂质双分子层在细胞或细胞器周围形成了一道疏水屏障, 将其与周围环境隔绝起来。
尽管有一些小分子可以直接渗透通过膜,但是大部分的亲水性化合物,如糖,氨基酸,离子,药物等等,都需要特异的转运蛋白的帮助来通过疏水屏障。
因此,转运蛋白在营养物质摄取,代谢产物释放以及信号转导等广泛的细胞活动中起着重要的作用。
大量疾病都与膜转运蛋白功能失常有关,转运蛋白是诸如抗抑郁剂,抗酸剂等大量药物的直接靶点。
研究主要集中在次级主动运输蛋白的工作机理上。
交替通路模型,被用来解释转运蛋白的工作机理,在这个模型中,转运蛋白至少采取两种构象来进行底物的装载及卸载:
一种向膜外开放,一种向膜内开放。有许多结构和生物物理学证据支持这个模型。
但是,仍有两个最有趣的基本问题没有解决。
第一,主动运输的能量偶联机制是什么?
第二,在转运过程中,是什么因素触发了转运蛋白的构象变化?使用基于结构的研究手段对次级主动运输蛋白进行研究,以期解决转运蛋白工作机理中的基本问题。
主要成就
2014年,颜宁率领的团队在世界上首次解析了人源葡萄糖转运蛋白GLUT1的三维晶体结构。
2015年进一步获得了具备更多构象的GLUT3结合底物和抑制剂的超高分辨率结构,从而清晰揭示了葡萄糖跨膜转运这一基本细胞过程的分子基础。
此外,她还对离子通道结构生物学领域做出重要贡献,解析了电压门控钠离子通道的晶体结构,最近又利用最新冷冻电镜技术获得了最大钙离子通道RyR1的高分辨率结构。
2015年进一步获得了具备更多构象的GLUT3结合底物和抑制剂的超高分辨率结构,从而清晰揭示了葡萄糖跨膜转运这一基本细胞过程的分子基础。
2016年9月-Science-关闭及开放构象的RyR2
2016年9月,颜宁教授研究组与加拿大卡尔加里大学陈穗荣研究组合作在《Science》(DOI:10.1126/science.aah5324)发表研究长文,揭示了已知分子量最大的离子通道Ryanodine受体RyR2亚型处于关闭和开放两种状态的三维电镜结构,探讨了RyR2的门控机制。
通过比较关闭和开放状态的两个结构,发现位于穿膜区域负责通透离子的通道有明显的变化:
在开放构象中,该通道发生扩张,从而使得钙离子能够顺利地从肌质网内部转移到细胞质中。通过对RyR2中每个相对独立的结构域的仔细比较和分析,认为中心结构域极有可能是引发RyR开放的关键,这一发现与之前有关RyR的功能研究结论相吻合。
另外,研究组还获得了分辨率为5.7埃的RyR1开放构象结构,并基于结构比对,初步分析了RyR1的门控机理,有关RyR1的成果已分别发表在《Nature》(Doi:10.1038/nature14063)和《Cell Research》(Doi:10.1038/cr.2016.89)上,有关Cav1.1的论文已分别发表于《Science》(DOI: 10.1126/science.aad2395)和《Nature》(Doi:10.1038/nature19321)杂志上。上述研究与最新的这篇研究论文极大地促进了人们对于兴奋-收缩偶联的理解。
2017年2月,真核生物电压门控钠离子通道的拓扑图和三维电镜结构
2017年2月,颜宁教授研究组在《科学》(Science, DOI: 10.1126/science.aal4326)在线发表了题为“Structure of a eukaryotic voltage-gated sodium channel at near atomic resolution”的研究长文,在世界上首次报道了真核生物电压门控钠离子通道。


 添加微信免费咨询
添加微信免费咨询
