氨基酸基因实变
【研究发现
#动辄大打出手或与基因突变有关#】
#研究称有攻击行为的人体内可能发生了基因突变#人群中,有人温文尔雅,有人暴虐无常。最新研究发现,那些有攻击行为的人,体内可能发生了基因突变。记者16日从南京大学获悉,该校医学院石云教授团队联合中外学者研究发现,编码大脑内谷氨酸受体GluA3的GRIA3基因突变,会引起人类攻击行为。该成果近日发表于国际精神病学领域旗舰学术期刊《分子精神病学》。
论文的共同通讯作者、南京大学医学院教授石云介绍,在过去的研究中,研究人员发现,将X染色体上编码谷氨酸受体GluA3的GRIA3基因从雄性小鼠体内敲除,小鼠会表现出强烈的攻击行为。所以,人类的同源基因GRIA3被学界普遍认为是人类攻击行为的易感基因之一。然而迄今为止没有直接证据表明GRIA3基因和人类的攻击行为相关。
“此次研究,我们在4个有突发攻击行为的欧洲患者中,发现他们携带有GRIA3基因的罕见变体,其中3个患者的GRIA3基因突变,导致GluA3谷氨酸受体发生G630R的氨基酸残基变化,另一个患者的基因突变导致了E787G的变化。功能检测表明,这两种突变都造成GluA3功能的丧失,相当于信号接收器坏了。”石云解释,也就是说,谷氨酸结合到突变的GluA3受体上,不能有效传递神经信号,因此初步判断人类GluA3的功能损坏会导致攻击行为,这和基因敲除小鼠的行为一致。
除了基因突变的病人,在普通人群中是否存在影响GluA3表达水平的遗传因素,诱发了攻击行为?
该论文的第一作者彭世笑说,研究人员在GRIA3基因的192个高频单核苷酸多态性位点中,发现rs3216834位点为连续的鸟苷酸重复序列,在人群中约78%的人带有9个鸟苷酸(9G),少数人带有7G、8G、10G和11G。实验表明,10G或11G的长的鸟苷酸重复序列会严重抑制GluA3受体的表达水平。
随后,研究者在294位男性暴力犯罪服刑人员中发现9人携带rs3216834-10G,另有1人携带11G,占总体的3.4%。而在没有犯罪记录的937位社区男性中,仅有2人携带10G(0.2%),并且没有发现11G的携带者。
“这说明rs3216834-10G、11G的携带者在暴力犯罪服刑人员中远高于正常对照人群,而10G和11G严重抑制GluA3的表达,增加了攻击行为的发生风险。”石云介绍。
为了进一步分析GluA3受体损坏或表达降低诱发攻击行为的神经机制,研究人员分析了GRIA3基因敲除小鼠,发现小鼠的内侧前额叶皮层的神经活动减低。而将前额叶皮层的GluA3补偿回去,可以显著缓解敲除小鼠的攻击行为。
“这意味着前额叶皮层异常的神经活动是攻击行为的关键神经环路机制。”石云认为,这项研究揭示了人类攻击行为新的遗传因素,这对于缓解攻击行为的药物研发,或可提供参考。(科技日报;艾塔)
我国新冠疫苗能否抵挡印度变异病毒
科研人员近日在印度奥里萨邦发现了73个新冠病毒毒株的新型变种,而就在几天前,日本科研人员也发现本国有多种变异的新冠病毒,其基因序列都与已知病毒亚型存在一定差异。这么多变异的新冠病毒对疫苗研发有什么影响?全球正在开发的多种疫苗,会不会无法预防病毒变种对人体的入侵?解放日报·上观新闻记者今天采访了上海交大医学院上海市免疫学研究所教授、上海市免疫学会副理事长王颖。
新冠病毒属于单链RNA(核糖核酸)病毒,其在宿主细胞内复制过程中由于RNA复制酶的保真性比较差,容易产生基因突变。不过目前看来,成熟的新冠病毒和SARS(严重急性呼吸综合征)病毒一样,其突变比例相对较低,甚至比常见的流感病毒要低。
“公众其实不必对新冠病毒的变异感到恐慌,这些变异造成目前研发疫苗失败的可能性是比较小的。”王颖分析说,因为从目前正在研发的疫苗类型来看,如果是灭活病毒疫苗,灭活病毒上所含有的表位数量众多,足以诱导免疫保护作用;如果采用的是病毒关键蛋白作为候选疫苗靶蛋白,那么需要对这些突变的位点做进一步分析,判断是否会让在研疫苗失效。
据介绍,全球用于疫苗研发的靶蛋白基本上是两种——S蛋白(刺突蛋白)和N蛋白(核衣壳蛋白),这些蛋白免疫人群后,可以诱导产生免疫保护。以S蛋白为例,新冠病毒正是通过这种蛋白质与人体细胞表面的ACE2(血管紧张素转化酶2)结合,从而入侵细胞。将靶标为S蛋白的新冠疫苗注射入人体后,免疫系统所产生的抗S蛋白的抗体就会阻断病毒进入宿主细胞。与此同时,抗S蛋白的免疫记忆可以使免疫过的个体再次感染后,迅速产生特异性抗体,从而阻止其进一步感染人体细胞,起到免疫保护作用。
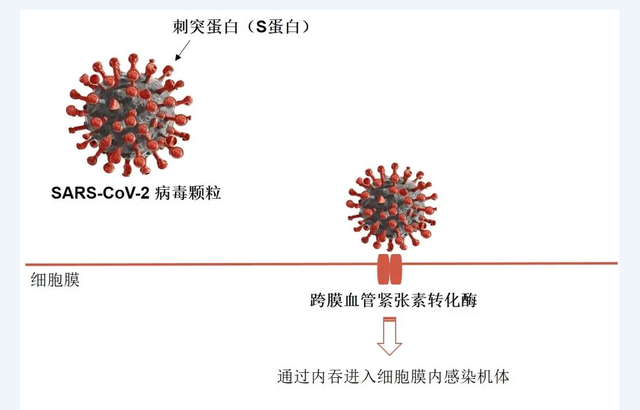
新冠病毒的S蛋白结合ACE2进入细胞。来源:君实生物
因此,新冠病毒基因突变如果没有造成编码蛋白的序列改变,就不会影响蛋白质诱导免疫,疫苗对突变病毒仍然是有效的。这类不影响编码氨基酸序列的基因突变叫作“无义突变”,对所编码的氨基酸有改变的突变则称为“有义突变”。王颖指出,“有义突变”还可以分为两种情况:一种是突变的氨基酸残基确实会影响蛋白质的免疫原性,即诱导特异性免疫反应;另一种情况是所发生突变的氨基酸并非位于主要表位区,对疫苗诱导产生免疫保护作用的影响不大。
因此,我们要辩证看待不断出现的新冠病毒变异研究结果。一方面,公众不必感到恐慌,因为目前在研新冠疫苗有足够多的位点可以产生免疫保护作用,更何况很多基因位点的突变不一定会让疫苗失效;另一方面,疫苗研发人员应关注新冠病毒变异的进展,探究这些基因突变的生物学意义,从而在疫苗研发中更加全面地把握候选疫苗的设计,确保可以诱导足够的免疫保护作用。

工作人员在国药集团中国生物新冠疫苗生产基地对灭活疫苗样品进行检测。新华社 发
王颖还指出,新冠病毒的变异可能具有一定地域性,各个地区的流行株可能存在不同的突变。这种地域性与人种的遗传背景有关,所以在印度发现的73个新型变种,在中国流行的可能性不大。这也与我国对新冠病毒疫情的防控有效性有关。这意味着,我国企业和科研机构开发的疫苗如果只是针对中国人群,那么主要关注的就是中国的流行株,而针对地域性突变株的研发可能是处于次要地位。
如果国产疫苗今后要出口到不同流行株的国家和地区,那么在疫苗研发中,就需要结合当地的流行株进行更有针对性的设计和研究,从而使疫苗对当地人群更为有效。
栏目主编:黄海华 文字编辑:俞陶然 题图来源:新华社
来源:作者:俞陶然
新冠病毒在印度、日本出现大量变种,在研疫苗会失效吗?,我国新冠疫苗能否抵挡印度变异病毒
肺鳞癌驱动基因获重大突破!
近日,《自然》发布:研究发现Nsd3是肺鳞癌的驱动基因之一。
NSD3驱动基因的机制是什么?
NSD3是组蛋白甲基转移酶,它通过给组蛋白H3的第36位氨基酸残基(H3K36)加两个甲基。如果NSD3水平增加,或者活性增强(例如T1232A变异),就会导致H3K36被过度甲基化。
咱们的基因组在细胞核里面都是缠绕在组蛋白上的,组蛋白的表观遗传学修饰(例如甲基化)会像“开关”一样,影响周边基因的表达。
而NSD3对H3K36的双甲基化修饰,会导致附近促癌基因的表达开关被打开,进而驱动肺鳞癌的进展。
但是目前临床上还没有NSD3抑制剂。
于是研究人员又在一个抑制剂库里筛选能抑制NSD3活性的药物。结果找到了4种溴原子抑制剂(BETi),小鼠实验表明,肺鳞癌确实对BETi敏感。
总而言之,这项研究成果确定了NSD3是肺鳞癌的驱动因素,为开发治疗肺鳞癌的靶向药物找到了合适的靶点。
全球每年有10万肺鳞癌患者体内存在8p11-12片段的扩增。另外,乳腺癌和其他恶性肿瘤中也常见8p11-12片段的扩增。
期待抑制NDS3相关药物早日进入临床研究阶段。
2 FGFRI纤维母细胞生长因子受体1 突变比例约4%(4/95),家族成员(包括FGFR1
FGFR2, FGFR3。FGFR4,是一种酪氨酸跨膜激酶受体。目前的临床试验已宣告失败。
2 PI3KCA基因的扩增远较突变常见,鳞癌可以高达33.1%,而腺瘤(6.2%)和小细胞瘤(4.7%)。
目前针对P3KCA基因突变的单药1期研究,包含肺癌、乳腺癌、结直肠癌等多种癌症。
4 DDR2盘状结构域受体(discoidin domain receptor 2, 中与在恶性黑色素宿中的常见突变V600E不同,值得进DDR2) 肺鳞痛DDR2突变率虽然不高,一项针对DDR2突变
肺鳞癌的临床研究已经结束人组(NCT 01491633 ),若研究发现肺鳞癌DDR2突变者对达沙替尼确实有效,将会对该亚型患者治疗带来突破性的研究进展。
5 MET基因扩增MET基因是一种编码HGFR蛋白原癌基因,是一种酪氨酸激酶膜受体,位于染色体721-q3,其与配体肝细胞生长因子( hepatocyte grow factor, HGF)均促进肿瘤发生1″, MET扩增可见于懒癌和腺癌、MET扩增与EGFR-TKI继发耐药有关,有报道认为是通过激话ERBB-3通路相关。尽管有报告认为在肺癌的扩增比例约3%-21%,但由于这是一种低水平的扩增,有学者认为突变的比率要低的多,估计肺鳞癌为1%,腺癌为2%
多项MET抑制剂正研究中,目前的研究一般均针对MET过表达的NSCLC.并非仅针对解癌。例如:克唑替尼是MET和ALK的双重抑制剂;多把点药物cabozantinib(XL184)以MET, RET和VEGF2为靶点0 Tivatinib(ARQ197)则是以MET为靶点的小分子药物,但有一定的细胞毒性1;而单克隆抗体MetMAb联合厄洛替尼治疗复治NSCLC。
6 BRAF BRAF是KRAS下游的丝氨酸/苏氨酸激酶,将sRAS鸟,苷三瞬酸连接到丝裂原活化蛋白激酶家族的下游蛋白,控制细胞增殖. RAF激商家族包括3个成员:ARAF, BRAF IRAFI (也称为CRAF) , BRA突变与酶活性增加相关,导致MAPK2HMAPK3组成型活化。
总之,目前EGFR-TKI的出现使得肺癌的靶向治疗成为现实,但针对肺鳞癌的靶向治疗研究进展相比腺癌明显比较慢,到目前为止尚未开展一项1期临床研究。目前也仅发现约40%肺鳞癌患者携带不同的驱动基因,还有相当多的聚动基因未被发现,肿瘤的形成是一个多基因参与的异常复杂的过程,现有的靶向药物如小分子激酶扣制剂大多是针对一个靶点而设计,因此只有发现更多的驱动基因,阐明基因之间的相互作用关系,开发出更多的靶向药物,联合应用或联合其它的治疗方式,才有可能收到理想的疗效。
总之,真正实现肺鳞癌的个体化靶向治疗仍然任重而道远。


 添加微信免费咨询
添加微信免费咨询
